

目前,很多研究都关注植物叶片的动态光合。这是因为:⑴Rubisco酶的激活速度决定能否增加光合原料(光、CO2等)的利用效率和植物的生产力(Carmo Silva et al., 2015);⑵气孔的响应速度与叶片的瞬时水分利用效率(WUEi)有关(Lawson and Blatt, 2014);⑶非光化学猝灭(NPQ)的驰豫速度关系到弱光条件下叶片的光合量子产率(Murchie and Nivogi 2011);⑷考虑动态光合,能够更准确地预测植被的潜在生产力(Kaiser et al., 2015)。
为了探究这些问题的答案,对叶片动态光合的特点和行为模式的研究至关重要。关于这方面的实验测量也是量化动态光合限制因子,并且估算这些限制因子如何受到环境条件影响的数据基础。
LI-6800高级光合荧光测量系统的Auto Program功能,可实现全自动稳态光响应曲线、稳态A-Ci曲线、动态A-Ci曲线、以及光合诱导气体交换的测量。
举个例子
以下所有数据均出自Taylor SH, Long SP. 2017 Slow induction of photosynthesis on shade to sun transitions in wheat may cost at least 21% of productivity. Phil. Trans. R.Soc. B 372:20160543.
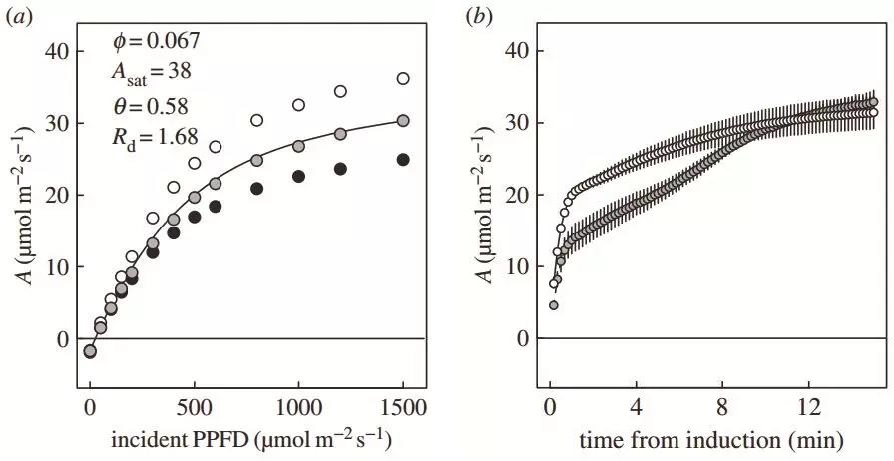
图1 a.三种实验材料的稳态光响应曲线;b. PPFD从50到1200 μmol/m2/s动态光合诱导曲线(空心圆+实线表示CO2浓度为100 μmol/mol时的光合诱导过程,实心圆+虚线表示CO2浓度为400 μmol/mol时的光合诱导过程)。
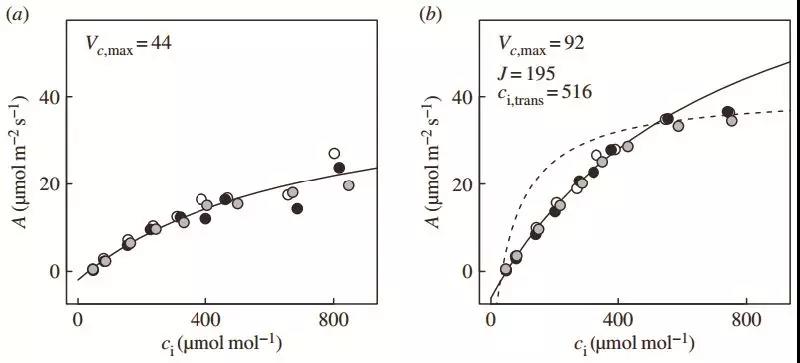
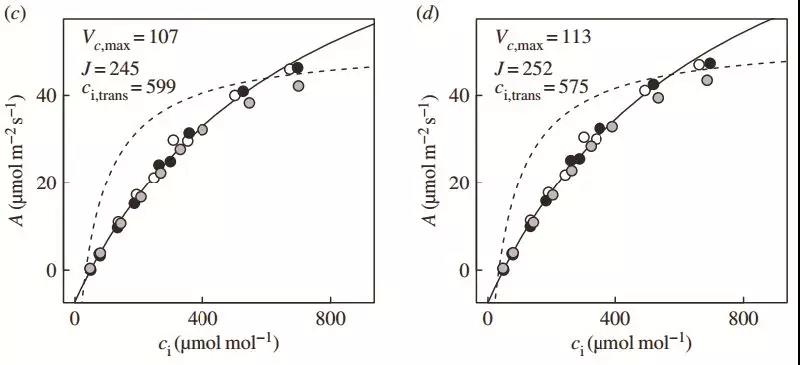
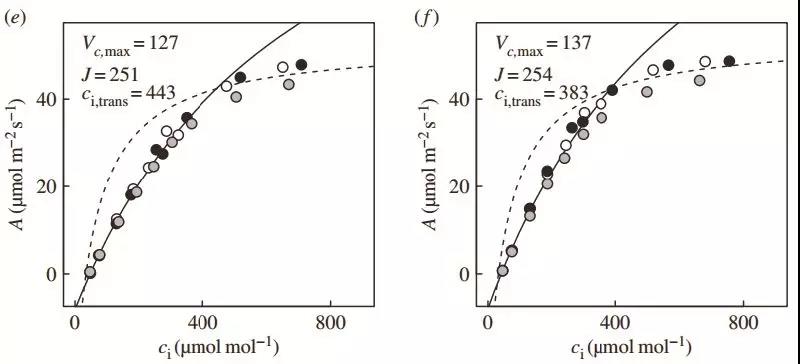
图2 从50 μmol/m2/s到1200μmol/m2/s的光合诱导过程中的动态A-Ci曲线 :a. 20 s;b. 1min;c. 2.5 min;d. 3 min;e. 4.5 min;f. 10 min。在光合诱导后的前40 s,植物光合主要受限于Ac,Vcmax也显著低于稳态光合。随着诱导进行,Vcmax和J均开始升高,且J升高要快于Vcmax。
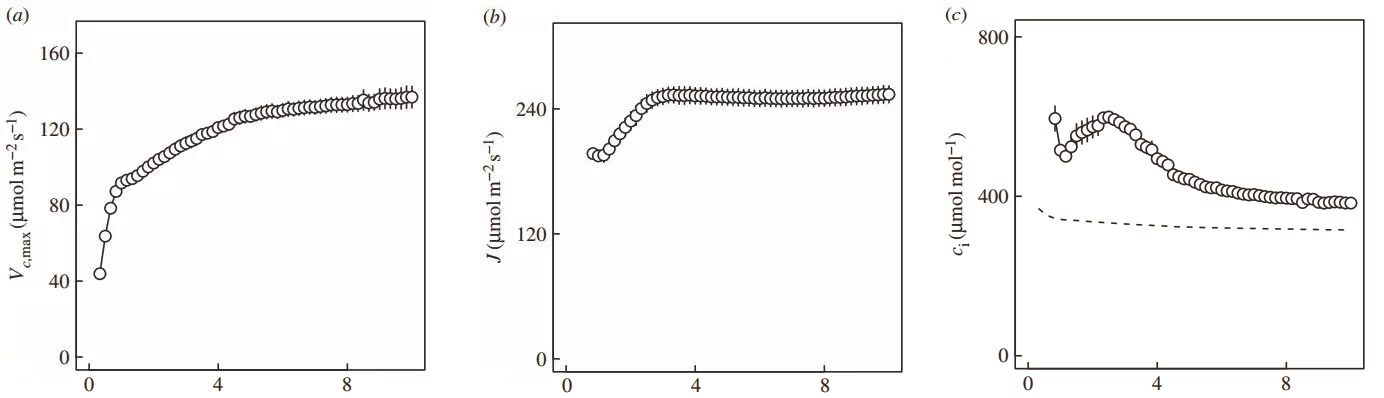
图3. 从50 μmol/m2/s到1200μmol/m2/s的10 min光合诱导过程中,光合限制的变化情况。a. Rubisco酶限制Vcmax;b. 电子传递限制J;c. Vcmax和J两种限制因素的拐点Ci。
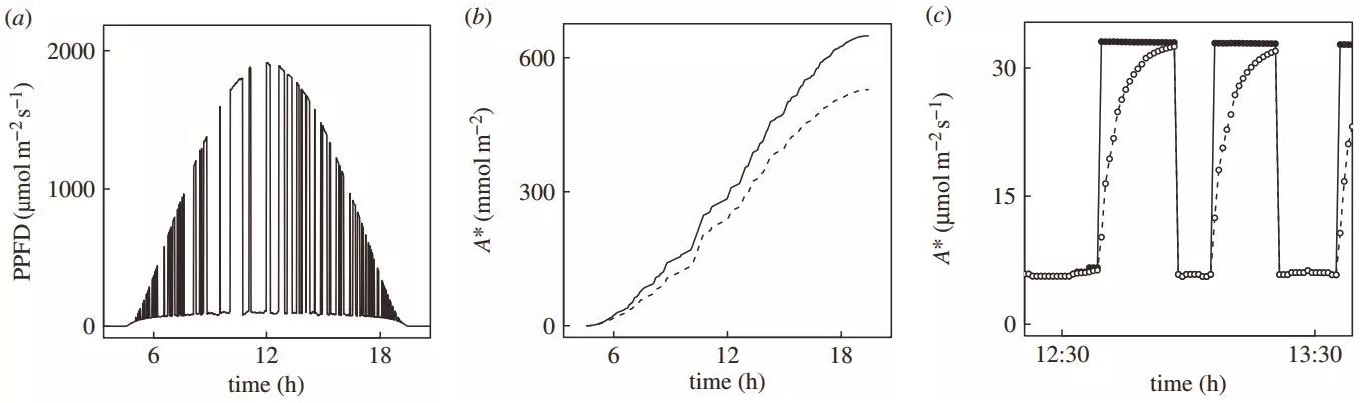
图4 a. 模拟叶片表面在晴朗的一天中所接受到的PPFD变化;b. 根据光响应曲线估算的一天总光合CO2累积量A*(实线) VS 考虑光合诱导引起的Rubisco再活化后估算的一天总光合CO2累积量A*(虚线);c. 正午时间段,无光合诱导滞后现象的光合速率(实心圆)和考虑Rubisco活化后的光合速率(空心圆)。
结论
通过对光合诱导过程的测量与分析,我们可以:⑴ 量化植物叶片在从遮阴到见光的光合诱导过程中的光合响应速度;⑵ 通过动态A-Ci曲线的测量,揭示环境因素对诱导适应速度的影响;⑶ 估算出由于慢速诱导适应所造成的的潜在产量损失。
参考文献
1. Taylor SH, Long SP. 2017 Slow induction of photosynthesis on shade to sun transitions in wheat may cost at least 21% of productivity. Phil. Trans. R. Soc. B 372:20160543.
2. Kaiser et al., 2017. Photosynthetic induction and its diffusional, carboxylation and electron transport processes as affected by CO2 partial pressure, tempera-ture, air humidity and blue irradiance. Annals of Botany 0: 1–15, 2016.
3. Woodrow IE, Mott KA. 1989 Rate limitation of nonsteady-state photosynthe-sis by ribulose-1,5-bisphosphate carboxylase in spinach. Aust. J. Plant Physiol. 16, 487–500.
4. Zhu XG, Ort DR, Whitmarsh J, Long SP. 2004 The slow reversibility of photosystem II thermal energy dissipation on transfer from high to low light may cause large losses in carbon gain by crop canopies: a theoretical analysis. J. Exp. Bot. 55, 1167–1175.
5. Zhang YP et al., 2018 Short-term Salt Stress Strongly Affects Dynamic Photosynthesis, but not Steady-State Photosynthesis, in Tomato (Solanum lycopersicum).Environmental and Experimental Botany. 149, 109-119.